
Dendropoma petraeum is a gastropod mollusk belonging to the Vermetidae family. This organism has very important abilities especially in the generation of bioconstructions between the mesolittoral and infralittoral belt. These structures are also called Trottoir a vermeti, that is, biogenic structures that generate a sort of reef.

“The Trottoir a vermeti is a marine bioconstruction that rises from the bottom towards the surface of the sea, gradually changing the local environment, both physically and ecologically” (Fagerstrom, 1987; Bianchi & Morri, 1996).
These reefs can be formed thanks to the fact that Dendropoma is a sessile organism, with a robust and irregularly shaped shell, colonial and gregarious, and above all, adapted to live in this area due to the presence of a horny operculum that closes the organism in the shell during periods of emergence, to then reopen it during periods of immersion.
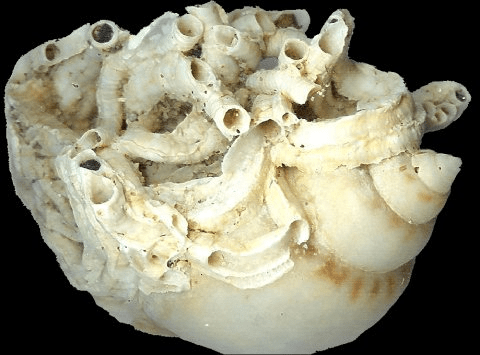
We are now aware that the formation of these structures increases biodiversity. Some of you may wonder why, and here’s the explanation. The complexity, structurality and three-dimensionality that generate these bioconstructions increase the surface area available for many organisms. Starting from the bacteria that begin to colonize the area, through what is called microbial biofilm, over time, bacterivorous organisms will be a source of food for other organisms and so on. Then think about colonization by algae, which attract herbivorous organisms that will be food for carnivorous ones. This doesn’t happen in all areas, or rather, it doesn’t happen in all areas with the same intensity. Because, if organisms can colonize a larger space due to the particularly accentuated three-dimensionality, especially compared to the rest of the coastal areas, then they will attract greater biodiversity.
For this reason, this species and these coastal constructions are protected by the Berne and Barcelona Conventions (conventions at European level aimed at protecting habitats and biodiversity).
Trottoir a vermeti is one of the most important habitats that we can find in the Mediterranean. In particular, from various molecular studies, it appears that there are 4 genetically and geographically distinct clades of Dendropoma petraeum, which are defined as cryptic, due to the fact that the organisms appear the same, even if at the genetic level both mitochondrial and nuclear are divergent. In fact, these organisms, although they have changed genetically, have had little or no ecological and morphological changes. The only obvious changes observed are those at the level of larval development.
For this we can define these 4 cryptic species, ecological equivalents between them, and with this we indicate organisms that occupy the same type of habitat and the same ecological role in the different areas in which they are found. In this case there was a type of allopatric speciation that generated the 4 different species, due to the absence of contact between them. Over time these species have differentiated and probably remain so because they are in different areas (We have already spoken of this type of speciation in The Barrier Between Atlantic & Pacific: The Speciation Process Of Ophioblennius spp.).
But what does all this have to do with the Messinian crisis? Here we are. Given the high genetic divergence between the species, these could have already differentiated before the Messinian crisis (5.9 – 5.3 Mya). However, these data go against the commonly accepted hypothesis that no stenoaline organism survived this period of high salinity and that therefore flora and fauna secondary recolonized the area.
The emergence of these data could make us think that some of these species may have survived this period, in the deeper areas or near the deltas of large rivers.
The analyzes that brought these data to light were carried out at 29 sites in the Mediterranean on an area covering 4000 km. These 4 lines of Dendropoma petraeum show a clear and delimited distribution from west to east: Western, Tyrrhenian-Sicilian, Ionian-Aegean and Levantine. This distinction in zones also allowed to observe the different moments in which the process of speciation and differentiation from the ancestral line began: the Tortonian (before the Messinian, during the Miocene). While, the most recent diversification events, date back to the period following the Messinian (between 3.75 and 0.52 Mya), are the differentiation events within the individual lineages already differentiated between the 4 areas.
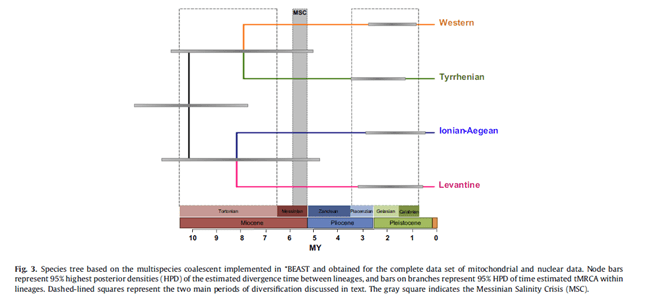
For this reason, the speciation process of these organisms took place in an estimated period ranging from 10 Mya to 4.5 Mya. During this period the Mediterranean, showed a high geographical diversity and a strong thermal zoning pattern (during the Messinian) due to the intense and long geological and oceanographic changes, that have always led to think that only euryhaline and brackish water species could survive (precisely because the Mediterranean was reduced to brackish or hypersaline lakes).
The time of divergence between these different species of Dendropoma suggests, that these organisms, survived during the Messinian almost certainly in the Tyrrhenian-Sicilian, Ionian-Aegean and Levantine basins. The same cannot be said with certainty for the western Mediterranean, which may have been replenished with species, following the reopening of the Strait of Gibraltar.
This is because, the drying process of the western Mediterranean has been almost complete, unlike the eastern Mediterranean which has maintained a significant water column. This leads us to assume, that probably, the saline conditions were not the same throughout the basin, and that therefore, there may have been “refugia” for stenoaline organisms able to survive, and to recover after this episodic event. The isolation in these refugia, obviously, reduced the size of the populations, increased the divergence between the various evolutionary lines and concluded the speciation process that had started millions of years earlier.
Sometimes it seems difficult to believe these scientific facts, because we feel weak, fragile, exposed to something that cannot be governed: the force of nature. Yet even in our absence, even without experimentation, repopulation, organism transplantation, even without our constant action, organisms resist.

Very often we think that the real ocean, the real treasure chest of life, is located offshore where we cannot see it, as if the ocean is constantly hiding something from us. I also thought this once. But then, in reality, I discovered that some of the richest habitats on the planet are located along the coast or almost dangerously close to it, we can think to mangroves, coral reefs, Trottoir a vermetidi (to cite some); these habitats allow the maintenance of the entire balance existing in the ocean.
Maria Bruno
M. Calvo et al. / Molecular Phylogenetics and Evolution 91 (2015) 17–26
